این مقاله در مورد زیست شناسی است، برای مشاهده زیر شاخه های آن جدول طبقه بندی زیست شناسی را ملاحظه کنید.
زیست شناسی
علم شناخت حیات است( این لغت از کلمه
یونانی بیاس به معنی زندگی و لوگاس یعنی دلیل منطقی تشکیل شده است). زیست شناسی به ویژگیها و
رفتارهای موجودات، چگونگی تشکیل
گونه ها و انواع موجودات و روابطی که آنها با هم دارند و به محیط زیست آنها مربوط می شود. زیست شناسی طیف گسترده ای از رشته های علمی که اغلب رشته های علمی مستقل بحساب می آیند را شامل می شود. روی هم رفته زیست شناسان حیات را از روی دامنه وسیعی از شاخصها مورد مطالعه قرار می دهند.
در مقیاس ذره ای و
مولکولی، زندگی مورد بررسی
زیست شناسی مولکولی،
زیست شیمی و علم
وراثت مولکولی است. در مقیاس
سلولی، مورد مطالعه زیست شناسی سلولی و در مقیاس های چند سلولی، مورد نظر
فیزیولوژی،
کالبد شناسی و
بافت شناسی است. زیست شناسی رشدی حیات را در مقیاس رشد و نمو اندام یک موجود مورد مطالعه قرار می دهد.
با بالا بردن مقیاس ها به بیش از یک موجود،
علم وراثت چگونگی عملکرد
وراثت بین والدین و فرزندان را مورد بررسی قرار می دهد.
رفتار شناسی جانوری رفتار گروهی بیش از یک موجود را مطالعه می کند. علم
وراثت جمعیتی میزان یک جمعیت کل را در در نظر دارد و علم
سیستماتیک شاخص چند گونه ای اجداد موجودات را بررسی می کند.
جمعیت های بهم وابسته ومحل سکونتشان در
بوم شناسی و زیست شناسی تکاملی مورد مطالعه قرار می گیرد. یک رشته نظری جدید
ستاره شناسی( یا زیست شناسى
گاز بى اثر
گزنون ) نام دارد که احتمالات وجود حیات در کرات دیگر غیر از زمین را مورد بررسی قرار می دهد.
زیست شناسی تنوع حیات را مورد برررسی قرار می دهد( در جهت عقربه های ساعت از سمت چپ در بالا) ای . کولی، درخت سرخس، بز کوهی و سوسک جالوت.
اصول زیست شناسی
گرچه زیست شناسی بر خلاف علم
فیزیک که معمولا سیستم های زیست شناختی را بر حسب اشیایی که تسلیم قوانین فیزیکی تغییر نا پذیر تشریح شده با
ریاضیات را توصیف نمی کند، با اینحال توسط بسیاری از اصول و مفاهیم اصلی توصیف می گردد که شامل: جامعیت، تکامل، تنوع، تسلسل، هم ایستایی و فعل و انفعالات می گردد.
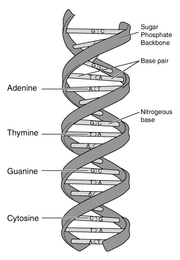 |
نمایش نمودار DNA در ابتدای
ماده تکوینی |
جامعیت: زیست شیمی، سلول ها و کد وراثتی
مقاله اصلی:
حیات واحد ها و فرهایند های رایج جامع بسیاری وجود دارد که برای گونه های مشخص
حیات ضروری می باشد. بعنوان مثال تمام گونه های حیات از
سلول هایی تشکیل شده اند، که در عوض، این سلول ها بر پایه یک سری زیست شیمی عمومی با زیر ساخت
کربن استوارند . تمام موجودات از طریق ماده وراثتی که بر پایه
نوکلئیک اسید دی ان ای استوار است از یک
کد وراثتیجامع استفاده می کنند.در مبحث رشد،موضوع فرایند های جامع نیز بیان می شود ، مثلا در بیشتر موجودات چند
یاخته اى قدمهای اولیه در رشد رویان مراحل ریخت شناسی مشابهی دارد و
ژنهای مشابهی را شامل می شود .
تکامل : هدف اصلی زیست شناسی
مقاله اصلی :
تکامل یکی از اهداف اصلی و سازمانده در زیست شناسی این است که تمام حیات از طریق یک فرایند
تکامل از یک
خاستگاه مشترک ناشی شده است . در واقع این یکی از عللی است که موجود زیستی تشابه قابل توجهی از واحدها و فرایند هایی که در بخش قبل تشریح شد را بروز می دهد .
چارلز داروین نظریه تکامل را بعنوان یک نظریه قابل دوام با برشمردن نیروی محرک آن بنا نهاد : نظریه انتخاب اصلح در
طبیعت . (
الفرد راسل والاس یکی از همکاران شخص پی برنده به این مفهوم شناخته می شود ) .
رانش وراثتی بعنوان یکی از شیوه های به اصطلاح ترکیب امروزی پذیرفته شده است .
تاریخچه تکاملی
گونه ها که گویای خصوصیات اجزای مختلفی است که از آن ناشی شده، به همراه رابطه شجره ای اش با دیگر گونه ها ،
تاریخ نژادی جانور یا گیاه نامیده می شود. دیدگاه های گوناگون زیادی در زیست شناسی اطلاعات مربوط به زیست شناسی را به وجود آورده است. این اطلاعات مقایسه های زنجیره های اسید دزوکسی ریبونوکلئیک که منجر به
زیست شناسی مولکولی و
ژنومیک می شود، و مقایسه های سنگواره ها با دیگر گونه های موجودات باستانی در علم
فسیل شناسی را شامل می شود. زیست شناسان روابط تکاملی را با روش های مختلف سامان داده و بررسی می کنند که این شیوه ها شامل
تکامل نژادی،
فنتیک، و
رده بندی جانداران بر حسب جد مشترک می شود. وقایع مهم در تکامل حیات، آنگونه که بتازگی زیست شناسان به آنها پی برده اند در این خط زمانی تکاملی بطور مختصر بیان شده است.
گوناگونی موجودات زنده
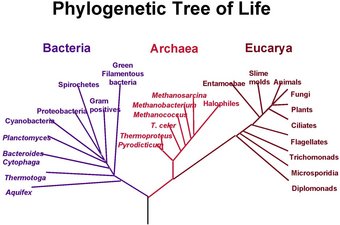 |
درخت روند تکامل تمام موجودات زنده مبنی بر اطلاعات
ژن RNA ،نمایش تفکیک سه قلمرو باکتری ها،جانوران اولیه
،موجودات چند سلولی شرح ابندایی توسط کارل ا.واس ،
ساختن درخت با دیگر ژن ها عموماَ شبیه هستند،
اگرچه آنها ممکن است در برخی مکان ها در شاخه های
نزدیک گروه های بسیار متفاوتی باشند ، احتمالاَ
زیرا بعلت تکامل سیع RNA نسبت دقیق از
سه قلمرو بحث هنوز وجود دارد. |
یک شجره تکامل نژادی از تمام موجودات زنده، مبنی بر داده های ژن اسیدیبونوکلئیک، نشانگر تفکیک سه مقوله باکتری، جانوران اولیه و موجوداتی که سلول تک هسته ای دارند می باشد. شجره هایی که با دیگر ژنها درست شده اند معمولا شبیه هم هستند، اگر چه ممکن است آنها برخی از گروههای زود دسته بندی شده را خیلی متفاوت از هم قرار دهند که احتمالا به سبب تکامل سریع اسید ریبونوکلئیک می باشد که در ابتدا توسط کارل ووس تشریح شد. روابط دقیق این سه مقوله هنوز مورد بررسی است.
علیرغم این وحدت اصولی، حیات نشانگر یک نوع گوناگونی زیاد حیرت انگیز در ساختار شناسی،
رفتار شناسی و
تاریخچه های حیات است. برای گلاویز شدن با این گوناگونی، زیست شناسان تلاش می کنند تا تمام موجودات را رده بندی کنند. رده بندی علمی باید منعکس کننده شجره های تکاملی ( شجره های وابسته به
تکامل نژادی ) موجودات مختلف باشد. این قبیل رده بندی ها قلمرو رشته های سیستماتیک و رده بندی جانداران را نشان می دهد. علم رده بندی جانوران ، جانداران را در رده هایی قرار می دهد که
تاکزا نامیده می شود، حال آنکه علم
سیستماتیک به دنبال روابط بین جانداران است. معمولا موجودات زنده به پنج سلسله تقسیم می شدند:
جلبک ها —
آغازیان –
قارچها —
گیاهان — جلبکها
بهرحال این پنج سیستم پنج سلسله ای اکنون دیگر بسیار قدیمی بحساب می آید. جایگزین های بسیار جدیدتر معمولا با
سیستم سه مقوله ای آغاز می شود:
جانوران اولیه ( در ابتداسلسله اصلی باکتری ها ) ،
باکتری ها ( در ابتدا ای یو باکتری) ،
موجودات چند سلولی این سه مقوله نشانگر این است که آیا سلولها هسته دارند یا خیر و همچنین منعکس کننده تفاوت ها در سطح خارجی سلولها است. همچنین یک سری از "انگل های" درون سلولی وجود دارند که به طور پیش رونده بر حسب فعالیت سوخت و سازی کمتر عمر می کنند:
ویروسها —
ویرید ها —
پریون ها
تسلسل: نژاد مشترک موجودات
مقاله اصلی:
نژاد مشترک اگر گروهی از جانداران با هم
جد مشترکی داشته باشند ، گفته می شود که این جانداران با هم نژاد مشترکی دارند. تمام
موجودات زنده روی
زمین از یک جد مشترک و یا
آبگیز ژن اجدادی زاده می شوند. گمان می رود آخرین جد همگانی ، یعنی ، جدیدترین جد مشترک تمام موجودات زنده
سه و نیم میلیون سال قبل بوجود آمده باشد ( به مبحث
منشا حیات رجوع کنید ) .
تصور اینکه منشأ " کل حیات از" یک تخم مرغ بوجود آمده است یکی از مفاهیم بنیادی زیست شناسی امروزی است ، بدین معنی که یک تسلسل نا شکسته از منشأ اولیه زندگی تا زمان حاضر وجود داشته است.تا
قرن نوزدهم معمولا اعتقاد بر این بود که گونه های حیات می توانند تحت شرایط خاصی خودبخود بوجود آیند. ( رجوع کنید به مبحث ایجاد
موجود زنده از مواد بی جان ). اصل عمومیت
کد وراثتی معمولا به عنوان ملاک تعیین کننده موافق نظریه جد مشترک جهانی ( یو سی دی ) برای تمام
باکتری ها،
موجودات اولیه و موجوداتی که یک هسته در سلولهایشان دارند مورد نظر زیست شناسان است. ( رجوع کنید به مبحث
سیستم سه مقوله ای ).
هم ایستایی: سازگار با دگر گونی
مقاله اصلی:
هم ایستایی هم ایستایی خاصیت یک
نظام باز برای تنظیم محیط درونی خودش است ، طوری که بوسیله چندین بار تطبیق همتراز حرکتی کنترل شده با ساز و کارهای قانونی بهم وابسته ، یک وضعیت ثابت را حفظ کند. تمام
موجودات زنده ، چه
تک سلولی و چه
چند سلولی هم ایستایی بروز می دهند . هم استایی در سطح سلولی می تواند با حفظ یک قدرت اسیدی ثابت خود را بروز دهد (
پی اچ ) ؛ در سطح موجود زنده حیوانات
خون گرم یک درجه حرارت ثابت درونی را حفظ می کنند ؛ و در سطح
بوم سازگان ، مثلا وقتی که میزان
دی اکسید کربن موجود در
جو افزایش می یابد ،
گیاهان قادر به رشد بهتری هستند و بنابراین دی اکسید کربن بیشتری از جو می زدایند. بافت ها و اندامها تیز می توانند هم ایستایی داشته باشند.
فعل و انفعالات : گروه ها و زیستگاه ها
همزیستی متقابل بین دلقک ماهی از جنس آمفیبریون است که در میان شاخک شقایق دریایی مناطق استوایی است وجود دارد. ماهی منطقه ای شقایق نعمان را از دست ماهی شقایق نعمان خوار محافظت می کند و در عوض شاخک حساس نیش زن شقایق نعمان از ماهی شقایق در برابر شکارچیان محافظت می کند. هر موجود زنده ای با دیگر مو جودات و محیط زیست خودش فعل و انفعال داخلی دارد. یکی از دلایلی که مطالعه سیستم های زیست شناختی را مشکل می سازد این است که فعل و انفعالات امکان پذیر مختلف بسیار زیادی با دیگر موجودات زنده و محیط زیست آنها وجود دارد. واکنش یک میکروب گیاهی بسیار ریز به یک شیب در حبه
قند به میزان واکنش آن به محیط زیست خود بهمان اندازه است که یک شیر هنگامی که در
دشت بی علف آفریقا در جستجوی غذاست نسبت به محیط زیست خود واکنش نشان می دهد. در گونه های خاصی
رفتارها می تواند دوستانه ،
تهاجمی ،
انگلی یا
همزیگری باشد. مسائل هنوز هم پیچیده تر می شود وقتی دو یا بیشتر گونه های مختلف در یک
بوم سازگان با هم فعل و انفعال داشته باشند و اکو سیستم یکی از شاخه های
بوم شناسی است
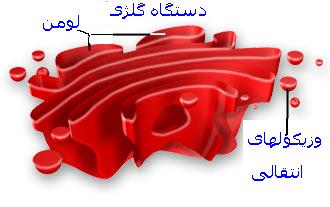 دستگاه گلژی
دستگاه گلژی 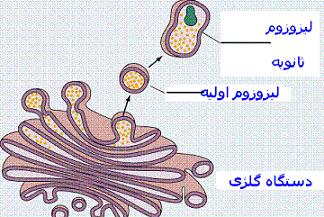
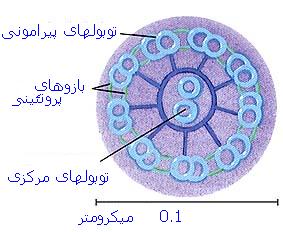
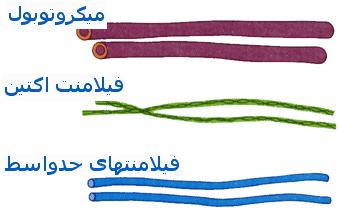



 متر) است.
متر) است.






